Expert medical al articolului

Noile publicații
Cauzele și patogeneza difteriei
Ultima examinare: 04.07.2025

Tot conținutul iLive este revizuit din punct de vedere medical sau verificat pentru a vă asigura cât mai multă precizie de fapt.
Avem linii directoare de aprovizionare stricte și legătura numai cu site-uri cu reputație media, instituții de cercetare academică și, ori de câte ori este posibil, studii medicale revizuite de experți. Rețineți că numerele din paranteze ([1], [2], etc.) sunt link-uri clickabile la aceste studii.
Dacă considerați că oricare dintre conținuturile noastre este inexactă, depășită sau îndoielnică, selectați-o și apăsați pe Ctrl + Enter.
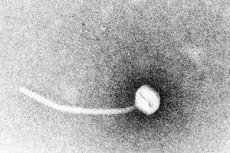
Difteria este o paradigmă a bolilor infecțioase toxigene. În 1883, Klebs a demonstrat că Corynebacterium diphtheriae era agentul cauzator al difteriei. Un an mai târziu, Loeffler a descoperit că organismul putea fi cultivat doar din cavitatea nazofaringiană și a propus că afectarea organelor interne se datora unei toxine solubile. Până în 1888, Roux și Yersin au arătat că animalele injectate cu filtrate sterile de C. diphtheriae au dezvoltat o patologie de organe imposibil de distins de cea a difteriei umane; acest lucru a demonstrat că o exotoxină puternică era principalul factor de virulență.
Difteria este cel mai adesea o infecție a tractului respirator superior, care provoacă febră, dureri în gât și stare generală de rău. O membrană groasă de fibrină, gri-verzuie, o pseudomembrană, se formează adesea la locul (locurile) infecției, ca urmare a efectelor combinate ale creșterii bacteriene, producției de toxine, necrozei țesuturilor subiacente și răspunsului imun al gazdei. Recunoașterea faptului că afectarea sistemică a organelor se datorează acțiunii toxinei difterice a condus la dezvoltarea atât a unei terapii eficiente pe bază de antitoxină pentru tratamentul infecției acute, cât și a unui vaccin toxoid extrem de eficient.
Deși imunizarea cu anatoxină a transformat difteria într-o boală rară în zonele în care standardele de sănătate publică impun vaccinarea, focarele de difterie apar încă în grupuri neimunizate și imunocompromise. În schimb, focare extinse de difterie care au atins proporții epidemice au avut loc în zonele în care programele de imunizare activă au fost suspendate.
Alte tipuri de corinebacterii
Pe lângă C. diphtheriae, C. ulcerans și C. pseudotuberculosis, C. pseudodiphtheriticum și C. xerosis pot provoca ocazional infecții nazofaringiene și cutanate. Ultimele două tulpini sunt recunoscute prin capacitatea lor de a produce pirazinamidază. În medicina veterinară, C. renale și C. kutscheri sunt agenți patogeni importanți, provocând pielonefrită la bovine și, respectiv, infecții latente la șoareci.
Cauze
Agentul cauzator al difteriei, Corynebacterium diphtheriae, este o tijă subțire, ușor curbată, cu îngroșări în formă de bâtă la capete, nemișcată; nu formează spori, capsule sau flageli și este gram-pozitivă.
Pe lângă toxină, corinebacteriile difterice produc în timpul proceselor lor vitale neuraminidază, hialuronidază, hemolizină, factori necrozanți și difuzi, care pot provoca necroză și lichefierea substanței principale a țesutului conjunctiv.
Pe baza capacității lor de a forma toxine, corinebacteriile difterice se împart în toxigene și non-toxigene.
Toxina difterică este o exotoxină bacteriană puternică care determină atât manifestările clinice generale, cât și cele locale ale bolii. Toxigenitatea este determinată genetic. Corinebacteriile netoxigene din difterie nu provoacă boala.
Conform caracteristicilor culturale și morfologice, toate corinebacteriile difterice sunt împărțite în 3 variante: gravis, mitis, intermedius. Nu există o dependență directă a severității bolii de varianta corinebacteriei difterice. Fiecare variantă conține atât tulpini toxigene, cât și non-toxigene. Corinebacteriile toxigene din toate variantele produc o toxină identică. [ 1 ]
Structură, clasificare și tipuri de antigene
Corynebacterium diphtheriae este o bacterie Gram-pozitivă, nemotilă, în formă de bâtă. Tulpinile care cresc în țesuturi sau în culturi in vitro mai vechi conțin pete fine în pereții celulari care permit decolorarea în timpul colorării Gram și duc la o reacție Gram variabilă. Culturile mai vechi conțin adesea granule metacromatice (polimetafosfat) care se colorează în albastru-violet cu albastru de metilen. Zaharurile din pereții celulari includ arabinoza, galactoza și manoza. În plus, se poate izola esterul toxic 6,6'-trehalozei, care conține acizi corinemicolic și corinemicolenic în concentrații echimolare. Se recunosc trei tipuri distincte de cultură: mitis, intermedius, gravis.
Majoritatea tulpinilor necesită acizi nicotinici și pantotenici pentru creștere; unele necesită și tiamină, biotină sau acid pimelic. Pentru o producție optimă de toxină difterică, mediul trebuie suplimentat cu aminoacizi și trebuie pus deoparte.
Încă din 1887, Loeffler a descris izolarea tulpinilor avirulente (nontoxigene) de C. diphtheriae, care erau imposibil de distins de tulpinile virulente (toxigene) izolate de la pacienți sănătoși. În prezent, se recunoaște că tulpinile avirulente de C. diphtheriae pot fi convertite într-un fenotip virulent în urma infecției și lizogenizării de către unul dintre mai mulți corinebacteriofagi distinși care poartă gena structurală pentru toxina difterică, tox. Conversia lizogenă a fenotipului avirulent în virulent poate avea loc atât in situ, cât și in vitro. Gena structurală pentru toxina difterică nu este esențială nici pentru corinebacteriofag, nici pentru C. diphtheriae. În ciuda acestei observații, nu s-a observat derivă genetică a toxinei difterice.
 [
[ Patogeneza
Punctele de intrare ale infecției sunt membranele mucoase ale orofaringelui, nasului, laringelui, mai rar mucoasa ochilor și a organelor genitale, precum și pielea deteriorată, suprafețele rănilor sau arsurilor, erupțiile cutanate de scutec, plăgile ombilicale nevindecate. La punctul de intrare, corynebacterium difteric se înmulțește și secretă exotoxină.
Exudatul bogat în fibrinogen este exudat și transformat în fibrină sub influența trombokinazei eliberate în timpul necrozei celulelor epiteliale. Se formează o peliculă fibrinoasă - semn caracteristic difteriei.
Transportul nazofaringian asimptomatic este frecvent în regiunile endemice pentru difterie. La persoanele susceptibile, tulpinile toxigene provoacă boala prin replicarea și secreția toxinei difterice în nazofaringe sau în leziunile cutanate. Leziunea difterică este adesea acoperită de o pseudomembrană compusă din fibrină, bacterii și celule inflamatorii. Toxina difterică poate fi scindată proteolitic în două fragmente: fragmentul A N-terminal (domeniu catalitic) și fragmentul B (domenii transmembranare și de legare a receptorului). Fragmentul A catalizează ADP-ribozilarea dependentă de NAD+ a factorului de elongație 2, inhibând astfel sinteza proteinelor în celulele eucariote. Fragmentul B se leagă de un receptor de suprafață celulară și facilitează livrarea fragmentului A în citosol.
Imunitatea protectoare implică răspunsul anticorpilor la toxina difterică după boala clinică sau la toxina difterică (toxină inactivată de formaldehidă) după imunizare.
Colonizare
Se cunosc puține lucruri despre factorii care mediază colonizarea C. diphtheriae. Cu toate acestea, este clar că alți factori decât producerea de toxină difterică contribuie la virulență. Studiile epidemiologice au arătat că un anumit lizotip poate persista într-o populație pentru perioade lungi de timp. Acesta poate fi ulterior înlocuit de un alt lizotip. Apariția și dominanța ulterioară a unui nou lizotip într-o populație sunt probabil legate de capacitatea sa de a coloniza și de a concura eficient în segmentul său de nișă ecologică nazofaringiană. Corynebacterium diphtheriae poate produce neuraminidază, care scindează acidul sialic de la suprafața celulară în componentele sale piruvat și acid N-acetilneuraminic. Factorul Cord (6,6'-di-O-micoil-α, α'-D-trehaloză) este o componentă de suprafață a C. diphtheriae, dar rolul său în colonizarea gazdei umane este neclar.
Producția de toxine difterice
Gena structurală a toxinei difterice, tox, aparține unei familii de corinebacteriofagi strâns înrudiți, dintre care β-fagul este cel mai bine studiat. Reglarea expresiei toxinei difterice este mediată de un represor activat de fier, DtxR, care este codificat de genomul C. diphtheriae. Expresia toxinei depinde de starea fiziologică a C. diphtheriae. În condițiile în care fierul devine substratul limitator al ratei de creștere, fierul se disociază de DtxR, gena toxicodendronului se activează, iar toxina difterică este sintetizată și secretată în mediul de cultură la rate maxime.
Toxina difterică este neobișnuit de puternică; pentru speciile sensibile (de exemplu, oameni, maimuțe, iepuri, cobai), o doză de doar 100 până la 150 ng/kg greutate corporală este letală. Toxina difterică constă dintr-un singur lanț polipeptidic de 535 de aminoacizi. Analizele biochimice, genetice și cu raze X arată că toxina constă din trei domenii structurale/funcționale:
- ADP-riboziltransferază N-terminală (domeniu catalitic);
- o regiune care facilitează livrarea domeniului catalitic prin membrana celulară (domeniu transmembranar);
- domeniul de legare a receptorilor celulelor eucariote.
În urma digestiei ușoare cu tripsină și a reducerii în condiții de denaturare, toxina difterică poate fi scindată specific la nivelul buclei sale sensibile la protează în două fragmente polipeptidice (A și B). Fragmentul A este componenta N-terminală de 21 kDa a toxinei și conține situsul catalitic pentru ADP-ribozilarea factorului de elongație 2 (EF-2).
Diagramă panglică a structurii cristaline cu raze X a toxinei difterice native monomerice. (modificată după Bennett MJ, Choe S, Eisenberg D: Domain swapping: Entangling alliances between proteins. Proc Natl Acad Sci, USA, 91: 3127, 1994). Sunt prezentate pozițiile relative ale domeniilor catalitice, transmembranare și de legare a receptorului. Toxina intactă poate fi scindată de proteazele de tip tripsină de la Arg190, Arg192 și/sau Arg193, care sunt situate în bucla sensibilă la protează (PSL). După reducerea punții disulfidice dintre Cys186 și Cys201, toxina poate fi separată în fragmentele A și B. Sunt prezentate capătul amino-terminal (N) și capătul carboxi-terminal (C) ale toxinei intacte. Diagrama tip bandă a fost creată folosind programul MOLESCRIPT.
Fragmentul C-terminal, fragmentul B, poartă domeniile transmembranar și de legare a receptorilor ale toxinei.
Intoxicația unei singure celule eucariote cu toxina difterică implică cel puțin patru etape distincte:
- legarea toxinei de receptorul său de pe suprafața celulară;
- gruparea receptorilor încărcați în fose acoperite și internalizarea toxinei prin endocitoză mediată de receptori; în urma acidificării veziculei endocitare de către o pompă de protoni asociată membranei, acționată de ATP,
- inserarea domeniului transmembranar în membrană și facilitarea eliberării domeniului catalitic în citosol și
- ADP-ribozilarea EF-2, rezultând o suprimare ireversibilă a sintezei proteinelor.
S-a demonstrat că o singură moleculă din domeniul catalitic administrată în citosol este suficientă pentru a fi letală pentru celulă.
Schema intoxicației difterice a unei celule eucariote susceptibile.
Toxina se leagă de receptorul său de la suprafața celulară și este internalizată prin endocitoză mediată de receptor; la acidificarea endosomului, domeniul transmembranar este inserat în membrana veziculei; domeniul catalitic este livrat citosolului, ducând la inhibarea sintezei proteinelor și la moartea celulară.
Epidemiologie
Înainte de imunizarea în masă a populației SUA cu anatoxină difterică, difteria era de obicei o boală a copilăriei. Un aspect remarcabil al imunizării în masă cu anatoxină difterică este că, pe măsură ce procentul din populație cu niveluri protectoare de imunitate la antitoxină (≥ 0,01 UI/ml) crește, frecvența izolării tulpinilor toxigene din populație scade. Astăzi, în Statele Unite, unde difteria clinică a dispărut aproape complet, izolarea tulpinilor toxigene de C. diphtheriae este rară. Deoarece infecția subclinică nu mai oferă o sursă de expunere la antigenul difteric și, dacă nu este stimulată, imunitatea la antitoxină scade, un procent mare de adulți (30 până la 60%) au niveluri de antitoxină sub nivelurile protectoare și sunt expuși riscului. În Statele Unite, Europa și Europa de Est, focarele recente de difterie au apărut în principal în rândul persoanelor care abuzează de alcool și/sau droguri. În cadrul acestui grup, purtătorii de C. diphtheriae toxigen au un nivel moderat ridicat de imunitate antitoxică. Recenta eșecare a măsurilor de sănătate publică din Rusia a dus la transformarea difteriei într-o epidemie. Până la sfârșitul anului 1994, în Rusia fuseseră raportate peste 80.000 de cazuri și peste 2.000 de decese.
Focarele focare de difterie sunt aproape întotdeauna asociate cu un purtător imun care s-a întors dintr-o regiune în care difteria este endemică. Într-adevăr, focarele recente de difterie clinică din Statele Unite și Europa au fost asociate cu călători care se întorc din Rusia și Europa de Est. Tulpinile toxigene de C. diphtheriae sunt transmise direct de la persoană la persoană prin picături aeriene. Se știe că tulpinile toxigene pot coloniza direct cavitatea nazofaringiană. În plus, gena toxigenă poate fi răspândită indirect prin eliberarea corinebacteriofagului toxigen și transformarea lizogenă a C. diphtheriae autohton netoxigen in situ. [ 5 ]
Pe lângă determinarea biotipului și lizotipului izolatelor de C. diphtheriae, tehnicile de biologie moleculară pot fi acum utilizate pentru a studia focarele de difterie. Modelele de digestie cu endonucleaze de restricție ale ADN-ului cromozomial de C. diphtheriae au fost utilizate pentru a studia focarele clinice, la fel ca și utilizarea secvențelor de inserție corinebacteriene clonate ca sondă genetică.
Testul Schick a fost utilizat timp de mulți ani pentru a evalua imunitatea la toxina difterică, deși acum a fost înlocuit în multe zone de testul serologic pentru anticorpi specifici împotriva toxinei difterice. În testul Schick, o cantitate mică de toxină difterică (aproximativ 0,8 ng în 0,2 ml) este injectată intradermic în antebraț (locul de testare) și 0,0124 μg de toxină difterică în 0,2 ml sunt injectate intradermic în locul de control. Citirile se fac la 48 și 96 de ore. Reacțiile cutanate nespecifice ating de obicei vârful la 48 de ore. La 96 de ore, o reacție eritematoasă cu posibilă necroză la locul de testare indică o imunitate antitoxică insuficientă pentru a neutraliza toxina (≤ 0,03 UI/ml). Inflamația atât în zona de testare, cât și în cea de control după 48 de ore indică o reacție de hipersensibilitate la preparatul antigenic.
Formulare
Corynebacterium diphtheriae infectează nazofaringele sau pielea. Tulpinile toxigene produc o exotoxină puternică ce poate provoca difterie. Simptomele difteriei includ faringită, febră și umflarea gâtului sau a zonei din jurul leziunii cutanate. Leziunile difterice sunt acoperite de o pseudomembrană. Toxina se răspândește prin fluxul sanguin către organele îndepărtate și poate provoca paralizie și insuficiență cardiacă congestivă. [ 6 ]
Există două tipuri de difterie clinică: nazofaringiană și cutanată. Simptomele difteriei faringiene variază de la faringită ușoară la hipoxie din cauza obstrucției căilor respiratorii de către o pseudomembrană. Afectarea ganglionilor limfatici cervicali poate provoca umflarea severă a gâtului (difterie de tip gât de taur), iar pacientul poate dezvolta febră (≥ 39°C). Leziunile cutanate în difteria cutanată sunt de obicei acoperite de o pseudomembrană gri-brună. Complicațiile sistemice care pun viața în pericol, în principal pierderea funcției motorii (de exemplu, dificultăți la înghițire) și insuficiența cardiacă congestivă, se pot dezvolta ca urmare a acțiunii toxinei difterice asupra neuronilor motori periferici și a miocardului.
Controla
Controlul difteriei depinde de imunizarea adecvată cu anatoxină difterică: toxină difterică inactivată de formaldehidă, care rămâne intactă din punct de vedere antigenic. Anatoxina se prepară prin incubarea toxinei difterice cu formaldehidă la 37°C în condiții alcaline. Imunizarea împotriva difteriei trebuie să înceapă în a doua lună de viață cu o serie de trei doze primare la intervale de 4 până la 8 săptămâni, urmate de o a patra doză la aproximativ 1 an după ultima doză primară. Anatoxina difterică este utilizată pe scară largă ca o componentă a vaccinului diftero-pertussis-tetanos (DPT). Studiile epidemiologice au arătat că imunizarea împotriva difteriei este eficientă în proporție de aproximativ 97%. Deși imunizarea în masă împotriva difteriei se practică în Statele Unite și Europa, iar ratele de imunizare a copiilor sunt adecvate, o mare parte din populația adultă poate avea titruri de anticorpi sub nivelurile protectoare. Adulții trebuie revaccinați cu anatoxină difterică la fiecare 10 ani. Într-adevăr, călătorilor în zone cu rate ridicate de difterie endemică (America Centrală și de Sud, Africa, Asia, Rusia și Europa de Est) ar trebui să li se administreze o imunizare de rapel cu anatoxină difterico-tetanică. În ultimii ani, utilizarea preparatelor de anatoxină înalt purificată pentru imunizare a redus la minimum reacțiile ocazionale severe de hipersensibilitate. Imunizarea de rapel cu anatoxină difterico-tetanică ar trebui administrată călătorilor în zone cu rate ridicate de difterie endemică (America Centrală și de Sud, Africa, Asia, Rusia și Europa de Est).
Deși antibioticele (cum ar fi penicilina și eritromicina) sunt utilizate ca parte a tratamentului pacienților cu difterie, imunizarea pasivă rapidă cu antitoxină difterică este cea mai eficientă în reducerea mortalității. Timpul lung de înjumătățire al antitoxinei specifice în fluxul sanguin este un factor important în asigurarea neutralizării eficiente a toxinei difterice; cu toate acestea, pentru a fi eficientă, antitoxina trebuie să reacționeze cu toxina înainte de a putea pătrunde în celulă.
Reproiectarea toxinei difterice pentru dezvoltarea de citotoxine specifice receptorilor eucarioți
Ingineria proteinelor este un domeniu nou și în rapidă dezvoltare în biologia moleculară; aceasta combină metodologiile ADN-ului recombinant și sinteza ADN-ului în fază solidă pentru a proiecta și construi gene himerice ale căror produse au proprietăți unice. Studiile relațiilor structură-funcție ale toxinei difterice au arătat clar că această toxină este o proteină cu trei domenii: catalitic, transmembranar și receptor. A fost posibilă înlocuirea genetică a domeniului nativ de legare a receptorului al toxinei difterice cu diverși hormoni polipeptidici și citokine (de exemplu, hormonul stimulator al α-melanocitelor [α-MSH], interleukina (IL) 2, IL-4, IL-6, IL-7, factorul de creștere epidermică). Proteinele himerice rezultate sau toxinele de fuziune combină specificitatea de legare a receptorului a citokinei cu domeniile transmembranar și catalitic ale toxinei. În fiecare caz, s-a demonstrat că toxinele de fuziune otrăvesc selectiv doar acele celule care poartă receptorul țintă corespunzător. Prima dintre aceste toxine de fuziune modificate genetic, DAB 389IL-2, este evaluată în prezent în studii clinice umane pentru tratamentul limfoamelor refractare și al bolilor autoimune în care celulele cu receptori IL-2 cu afinitate ridicată joacă un rol important în patogeneză.[ 7 ] Administrarea de DAB 389 IL-2 s-a dovedit a fi sigură, bine tolerată și capabilă să inducă o remisie durabilă a bolii, fără efecte secundare grave. Este probabil ca toxinele de fuziune pe bază de toxină difterică să devină noi agenți biologici importanți pentru tratamentul anumitor tumori sau boli în care pot fi vizați anumiți receptori de suprafață celulară.